
Caratteristiche salienti dei virus
Innanzitutto i virus sono caratterizzati da delle dimensioni estremamente ridotte, siamo nell'ordine del nanometro, i virus più piccoli come i poliovirus hanno un diametro di circa 20 nm. I virus più grandi arrivano ad avere una dimensione più grande di circa 500 nm. Siamo al di sotto dei limiti di risoluzione per il microscopio ottico, di conseguenza, se noi vogliamo osservare un virus dobbiamo ricorrere a un microscopio elettronico.
Per farvi entrare in questo ordine di grandezza, 1 nm corrisponde a 10⁻⁹ metri, quindi delle misure estremamente ridotte.
Non c'è assolutamente nessuna proporzione tra una struttura cosi semplice e il guaio che possono causare. Il più piccolo, di informazione genetica, tra i virus umani è l'epatite B, a dispetto di una pochezza incredibile di informazione genetica per malattie epatiche associate all'infezione di questo virus: muoiono 1 milione di persone all'anno.
In questa diapositiva, un virus è 10⁷ più piccolo di un pallone, il quale è a sua volta 10⁷ più piccolo del globo terrestre. Quindi, prima caratteristica saliente e importante per poter capire tutte le successive caratteristiche di questi virus sono le loro dimensioni estremamente ridotte.
Ma forse la caratteristica più saliente è questa, i virus sono dipendenti da una cellula vivente per la loro replicazione: in ambiente extracellulare un virus che è quiescente, deve, trovando specifici recettori attaccati alla cellula, entrarvi dentro e riuscire così a replicarsi.
Perché un virus dipende dalla cellula ospite?
Perché, innanzitutto manca di quelli che sono gli elementi base necessari per la replicazione e che sono invece presenti in tutte le cellule che, al contrario, sono capaci di farlo.
Il virus manca di quegli enzimi che sono indispensabili per la sintesi, di tutti quei precursori che partecipano alle sintesi macromolecolari, i virus non sanno sintetizzare nucleotidi, non sanno sintetizzare carboidrati, non sanno sintetizzare lipidi ne tanto meno amminoacidi, ecco che questo è un motivo fondamentale per il quale il virus dice alla cellula “aiutami vienimi incontro perché certo io non posso affrontare una sintesi proteica se non so prepararmi tutti quei mattoni che sono indispensabili per affrontare un simile processo”. Sono privi di tutti quei sistemi enzimatici che generano, normalmente sottoforma di ATP, energia chimica utilizzabile, ecco che ancora una volta il virus ricorre alla cellula per produrre tutta quella energia che serve per lo svolgimento delle sintesi virus specifiche. Manca di ribosomi, di RNA transfer, come pure di tutto quel macchinario enzimatico che è indispensabile perché all'interno di una cellula si sintetizzi una proteina. Manca di membrane, che sono la sede di tutte le sintesi macromolecolari normalmente realizzate all'interno di una cellula. Tutti e 5 motivi fondamentali che ci spiegano perché un virus all'esterno di una cellula è incapace di andare incontro a qualsiasi tipo di sintesi macromolecolare.Un'altra caratteristica è che un virus penetrato in una cellula, che vuole intraprendere un proprio processo replicativo, perde la propria integrità. Sapete dalla microbiologia, quando vi ho detto parassiti intracellulari obbligati che questa non è una caratteristica unica dei virus, ci sono anche cellule batteriche come per esempio le clamidie che replicano solo all'interno di una cellula. Ma che caratteristiche hanno questi procarioti? Vivono all'interno di una cellula ma sono capaci di una crescita alla quale segue una normale divisione in due cellule figlie, sono microrganismi che nel loro processo di crescita che culmina in un processo di divisione, non perdono la loro integrità.
Ben diverso è il comportamento dei virus: subito dopo che un virus è riuscito a penetrare all'interno di una cellula, o proprio durante questo processo di penetrazione, il virus in un processo chiamato UNCOATING si spoglia, si ha separazione tra il genoma e quelli che sono i rivestimenti superficiali del virus. Ecco che, a questo punto, il genoma virale diventa la centralina che dirige la replicazione e la formazione di una nuova progenie virale.
E che sintesi dovrà dirigere, questo genoma, per arrivare alla formazione di nuove particelle virali?
Deve fare due cose: tanti nuovi genomi e tante nuove proteine strutturali che, in una tappa, che poi è la tappa finale del ciclo replicativo si assemblano insieme per formare nuove particelle virali.

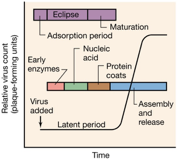
Lo vediamo meglio nella successiva diapositiva, a sinistra abbiamo una tipica curva di crescita di una cellula batterica: inizia con un periodo di latenza nella quale la cellula batterica si fornisce tutte le attività enzimatiche che sono indispensabili alla sua crescita e successiva divisione. Una volta che questo è realizzato abbiamo, una fase esponenziale, nella quale le cellule batteriche replicano, fino a che, esauriti i nutrimenti a disposizione si entra in una fase stazionaria che precede l’ultima fase che è quella di morte. In tutte queste fasi la cellula batterica è una cellula completa integra che, quando cresce e si divide nella fase esponenziale da una cellula se ne originano altre due.
Completamente diverso il comportamento replicativo da parte di un virus, ovviamente tutte le volte che voglio coltivare un virus in laboratorio devo avere a disposizione cellule. Nessun tipo di terreno ha la possibilità di supportare la replicazione di questi microrganismi essendo intracellulari obbligati.
Cosa succede quando io infetto delle cellule con un virus?
Lascio passare un certo periodo, pochi minuti, frammento quelle cellule, le apro, per vedere quello che c’è dentro, non trovo più particelle virali complete. Il virus a seguito dell’uncoating si è svestito, vedo che questo periodo dura un certo tempo prima che si possano di nuovo riscontrare in quella cellula particelle virali complete.
Che cosa fa il virus in questo periodo di latenza?
Vuole fare tanti nuovi genomi e sintetizzare proteine capsidiche, poi le acchiappa le mette insieme e si formano nuove particelle virali. Cosa vuole fare? Prima di tutto deve pensare a procurarsi quegli enzimi che gli servono per duplicare il proprio dna, e anche quelle attività enzimatiche che gli consentono di contrattare la cellula. Cosa vuol dire contrattare? Il virus entra all’interno di una cellula e dice, a questo punto la centralina di questa cellula che detta ordini è il mio genoma. Molto spesso un virus deve aver evoluto delle modalità che gli consentono di prendere il sopravvento su quelle che sono le sintesi molecolari della cellula ed è quello che fa con quelle che sono le proteine virali precoci. Dopo di che pensa a replicare il proprio genoma, quando ha replicato il proprio genoma sullo stampo di questi nuovi genomi si sintetizza le proprie proteine capsidiche ed eventuali altre proteine strutturali. A questo punto ha a disposizione nuovi genomi, nuove proteine strutturali ed ecco che il periodo di latenza finisce, avviene l’assemblaggio, e nuove particelle virali, potranno, se noi tramutiamo quella cellula prima che il virus sia fuoriuscito in ambiente extracellulare, ritroviamo nuovamente particelle virali.
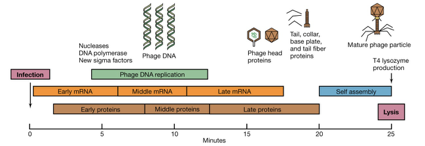
Osserviamo le fasi dell’infezione del fago T4. È un batteriofago, ossia un virus che ha come ospite una cellula batterica, che realizza il proprio ciclo replicativo in 25 minuti ma lo fa con una intelligenza che lascia sbalorditi, si è organizzato per fare ogni cosa al momento opportuno. È importante sottolineare come un microrganismo cosi piccolo possa, passo dopo passo, fare tutto quello che gli serve e, una volta per esempio, quando una attività enzimatica ha cessato di essergli utile ne stoppa la produzione.
Il virus è utilizzato, quindi, per infettare la cellula ospite. Si tratta di cellule di E.coli e cosa fa nella fase di latenza, nella quale se noi rompiamo quell’E.coli non vediamo più batteriofagi integri? Penetra nella cellula solo il genoma, il quale sintetizza prima degli Rna messaggeri che chiamiamo precoci, che servono al virus per duplicare il Dna genomico. Dopodiché con una serie di messaggeri che portano alla sintesi di proteine intermedie, è pronto per duplicare il proprio Dna. Una volta che, tanti nuovi genomi si sono formati, questi fungono da stampo per la formazione di messaggeri tardivi, che portano l’informazione per la sintesi delle proteine strutturali. Una volta che avremmo tanti nuovi genomi e tante nuove proteine strutturali segue l’ultima fase, l’assemblaggio, alla quale seguirà lo scoppio, perché il virus produce lisozima, dell’E.coli, e il riversarsi in ambiente extracellulare di una nuova progenie virale.
Il virus è utilizzato, quindi, per infettare la cellula ospite. Si tratta di cellule di E.coli e cosa fa nella fase di latenza, nella quale se noi rompiamo quell’E.coli non vediamo più batteriofagi integri? Penetra nella cellula solo il genoma, il quale sintetizza prima degli Rna messaggeri che chiamiamo precoci, che servono al virus per duplicare il Dna genomico. Dopodiché con una serie di messaggeri che portano alla sintesi di proteine intermedie, è pronto per duplicare il proprio Dna. Una volta che, tanti nuovi genomi si sono formati, questi fungono da stampo per la formazione di messaggeri tardivi, che portano l’informazione per la sintesi delle proteine strutturali. Una volta che avremmo tanti nuovi genomi e tante nuove proteine strutturali segue l’ultima fase, l’assemblaggio, alla quale seguirà lo scoppio, perché il virus produce lisozima, dell’E.coli, e il riversarsi in ambiente extracellulare di una nuova progenie virale.
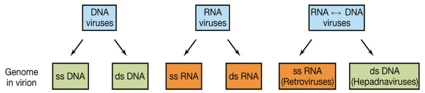
Abbiamo parlato di dimensioni dell'ordine dei nanometri, e sottolineato la peculiarità di un ciclo replicativo durante il quale un virus perde la propria integrità, altra cosa importante è che i virus hanno un genoma costituito o da Dna o da Rna mai entrambe queste molecole presenti contemporaneamente.
Cosa significa questo? Noi sappiamo bene che il flusso di informazione genetica va da Dna a Dna, da Dna a Rna, dall'Rna alle proteine, ciò significa: un virus con genoma costituito da Dna, una volta che è all'interno della cellula, può inserirsi tranquillamente in quello che è il normale flusso di informazione della cellula. I virus con genoma costituito da Rna si trovano a dover affrontare un grosso problema per la loro espressione. La cellula ospite, per formare dei messaggeri, utilizza un Rna polimerasi Dna dipendente, un virus che ha un genoma a Rna non può utilizzare questa attività enzimatica. La cellula non gli può venire incontro perché, essendo il flusso dell'informazione Dna-Rna-Proteine, la cellula non utilizza una Rna polimerasi Rna dipendente. Il virus codifica una Rna polimerasi Rna dipendente che, consente al genoma di potersi esprimere.
(Quando studiate dovete sempre chiedervi: se il genoma del virus sia a Rna o Dna e quanto Dna o Rna abbia.)
I virus a Dna (deossiribo virus) e i virus a Rna (ribovirus) possono avere un genoma a singola catena o a doppia catena. Una classe particolare di virus, i Retrovirus e i virus dell'epatite B, hanno l'informazione genetica contenuta in Dna o Rna ma in diversi momenti del ciclo di replicazione virale. L'esempio classico di retrovirus è l'HIV: il genoma è costituito da un Rna lineare, possiede una particolare attività enzimatica che è la trascrittasi inversa, cosi il genoma a Rna, durante il ciclo di replicazione di questo virus, viene convertito in Dna che poi va a integrarsi nel cromosoma della cellula ospite. Il virus dell'epatite B fa esattamente il contrario: ha un genoma costituito da Dna, nella particella virale completa e infettante ma, nel corso del ciclo replicativo, il virus ha scelto come via evolutiva quella di trasferire la sua informazione dal Dna all'Rna. Dopo aver visto che genoma contiene quel virus andiamo a vedere quanta informazione questo virus contiene. I Dna virus hanno un'informazione che va da 3,2 a 200 Kbp quindi enorme varietà. Gli Rna virus hanno un'informazione che va da 1,7 a 27 Kbp.
(Quando studiate dovete sempre chiedervi: se il genoma del virus sia a Rna o Dna e quanto Dna o Rna abbia.)
I virus a Dna (deossiribo virus) e i virus a Rna (ribovirus) possono avere un genoma a singola catena o a doppia catena. Una classe particolare di virus, i Retrovirus e i virus dell'epatite B, hanno l'informazione genetica contenuta in Dna o Rna ma in diversi momenti del ciclo di replicazione virale. L'esempio classico di retrovirus è l'HIV: il genoma è costituito da un Rna lineare, possiede una particolare attività enzimatica che è la trascrittasi inversa, cosi il genoma a Rna, durante il ciclo di replicazione di questo virus, viene convertito in Dna che poi va a integrarsi nel cromosoma della cellula ospite. Il virus dell'epatite B fa esattamente il contrario: ha un genoma costituito da Dna, nella particella virale completa e infettante ma, nel corso del ciclo replicativo, il virus ha scelto come via evolutiva quella di trasferire la sua informazione dal Dna all'Rna. Dopo aver visto che genoma contiene quel virus andiamo a vedere quanta informazione questo virus contiene. I Dna virus hanno un'informazione che va da 3,2 a 200 Kbp quindi enorme varietà. Gli Rna virus hanno un'informazione che va da 1,7 a 27 Kbp.
Perché è importante riflettere sulla quantità di informazione che un virus contiene?
Un virus molto piccolo, 20 nm di diametro, ha un'informazione genetica di soltanto 2000 nucleotidi. È un virus che ha un'informazione genetica talmente scarsa che, soltanto sintetizzando due proteine è in grado di terminare la propria replicazione. All'estremo opposto, un virus grande (esempio del mimivirus), circa 500 nm di diametro, 1,2 milioni di nucleotidi, ha la possibilità di sintetizzare 1200 proteine.
È importante avere questa informazione perché, immediatamente vi dice che le strategie replicative di questo piccolo virus, paragonate rispetto a quello più grande, saranno profondamente, tra loro, differenti.
Qui, stiamo analizzando un virus, mi viene in mente il fago MS2 che, per realizzare la propria sintesi è dipendente, in maniera massiva dalla cellula ospite. Questo virus, più ricco di informazione genetica, che scende in campo con molte proteine virali, sarà molto meno dipendente e potrà arrivare, per esempio, ad aumentare il proprio genoma in maniera indipendente dalla cellula ospite.
È importante avere questa informazione perché, immediatamente vi dice che le strategie replicative di questo piccolo virus, paragonate rispetto a quello più grande, saranno profondamente, tra loro, differenti.
Qui, stiamo analizzando un virus, mi viene in mente il fago MS2 che, per realizzare la propria sintesi è dipendente, in maniera massiva dalla cellula ospite. Questo virus, più ricco di informazione genetica, che scende in campo con molte proteine virali, sarà molto meno dipendente e potrà arrivare, per esempio, ad aumentare il proprio genoma in maniera indipendente dalla cellula ospite.
Cosa voglio dire con duplicare in maniera autonoma il proprio genoma?
Non soltanto per la sua polimerasi ma per tutte quelle attività enzimatiche che gli servono per duplicare il proprio genoma.
Quindi è importante la valutazione della quantità del Dna o Rna, e ricordiamo il problema che devono risolvere gli Rna virus: hanno un template di Rna, entra una molecola di Rna nel citoplasma di una cellula, la cellula non gli offre nessuna Rna polimerasi Rna dipendente, però il virus non ne può fare a meno. Non ne può fare a meno per due motivi: questa attività enzimatica gli serve per copiare l'Rna genomico e fare tanti nuovi Rna genomici, in parole più corrette deve duplicare il proprio genoma, come pure gli è indispensabile per sintetizzare i propri Rna messaggeri. Abbiamo visto come, la stragrande maggioranza dei virus codifica per una propria Rna polimerasi Rna dipendente che è in grado di consentire al virus entrambe queste funzioni: duplicare il proprio genoma e sintetizzare i propri Rna messaggeri indispensabili alla realizzazione di un ciclo replicativo.
Quindi è importante la valutazione della quantità del Dna o Rna, e ricordiamo il problema che devono risolvere gli Rna virus: hanno un template di Rna, entra una molecola di Rna nel citoplasma di una cellula, la cellula non gli offre nessuna Rna polimerasi Rna dipendente, però il virus non ne può fare a meno. Non ne può fare a meno per due motivi: questa attività enzimatica gli serve per copiare l'Rna genomico e fare tanti nuovi Rna genomici, in parole più corrette deve duplicare il proprio genoma, come pure gli è indispensabile per sintetizzare i propri Rna messaggeri. Abbiamo visto come, la stragrande maggioranza dei virus codifica per una propria Rna polimerasi Rna dipendente che è in grado di consentire al virus entrambe queste funzioni: duplicare il proprio genoma e sintetizzare i propri Rna messaggeri indispensabili alla realizzazione di un ciclo replicativo.
È importante sapere, all'inizio dello studio di un virus, quantità ecc, anche perché ci da un'idea di quella che è la capacità di questo virus di evolversi.
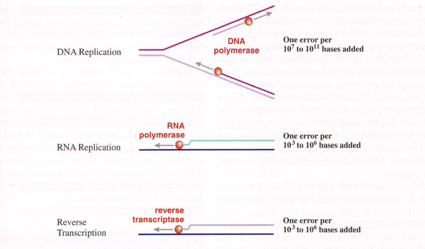
Perché abbiamo un diverso ritmo di capacità evolutiva tra gli Rna, che sono rapidissimi e i Dna che ovviamente hanno un ritmo ben inferiore?
Dipende, dalla serietà con cui lavorano gli enzimi che sono preposti alla replicazione degli acidi nucleici.
I Dna virus duplicano ricorrendo a una Dna polimerasi Dna dipendente. Questa è un'attività enzimatica precisa, che svolge bene il proprio lavoro che addirittura, che se compie un errore, è capace di fermarsi e tornare indietro per correggere. A dispetto di questo è anche vero che, la Dna polimerasi, ogni 10⁷ - 10^11 basi che aggiunge compie un errore. Si ha l'insorgenza di mutazioni e queste mutazioni sono uno degli avvenimenti principali che concorrono al processo evolutivo dei virus.
Come funziona una Rna polimerasi Rna dipendente?
È priva della possibilità di correggere errori, è un enzima paragonato alla Dna polimerasi che, in maniera più semplicistica possiamo dire che è più arrafazionale(?). Arriva a compiere un errore ogni 10^3 - 10⁵ paia di basi. Ecco che, i virus a Rna, a seguito di questo tasso mutazionale così elevato mutano con estrema velocità (con conseguenze molto importanti per l'ospite).
Tenete presente che questa situazione ha delle ripercussioni molto importanti sotto vari aspetti.
Anche la trascrittasi inversa, che i retrovirus utilizzano per convertire l'informazione genica da Rna a Dna, che poi viene integrato nel cromosoma ospite, anche questo, come la Rna polimerasi Rna dipendente, è un enzima piuttosto impreciso: ha lo stesso valore per quanto riguarda gli errori durante il funzionamento, un errore ogni 10^3 - 10⁶ basi aggiunte. Il genoma dell'HIV, che nel corso della propria replicazione utilizza una trascrittasi inversa con questo tasso mutazionale, ha un genoma costituito da 10⁴ ribonucleotidi. Ogni volta che la trascrittasi inversa legge, o meglio copia questo genoma a Rna per formare una molecola di Dna, inserisce per lo meno un errore. Se noi pensiamo che in un individuo infetto da HIV, si può arrivare ad avere la produzione di 10^11 particelle di HIV ci rendiamo conto dei mutanti che insorgono in un solo ciclo di replicazione nelle 24 ore. Questo giustifica la difficoltà ad ottenere un vaccino, lo stesso è vero per un farmaco.
I Dna virus duplicano ricorrendo a una Dna polimerasi Dna dipendente. Questa è un'attività enzimatica precisa, che svolge bene il proprio lavoro che addirittura, che se compie un errore, è capace di fermarsi e tornare indietro per correggere. A dispetto di questo è anche vero che, la Dna polimerasi, ogni 10⁷ - 10^11 basi che aggiunge compie un errore. Si ha l'insorgenza di mutazioni e queste mutazioni sono uno degli avvenimenti principali che concorrono al processo evolutivo dei virus.
Come funziona una Rna polimerasi Rna dipendente?
È priva della possibilità di correggere errori, è un enzima paragonato alla Dna polimerasi che, in maniera più semplicistica possiamo dire che è più arrafazionale(?). Arriva a compiere un errore ogni 10^3 - 10⁵ paia di basi. Ecco che, i virus a Rna, a seguito di questo tasso mutazionale così elevato mutano con estrema velocità (con conseguenze molto importanti per l'ospite).
Tenete presente che questa situazione ha delle ripercussioni molto importanti sotto vari aspetti.
Anche la trascrittasi inversa, che i retrovirus utilizzano per convertire l'informazione genica da Rna a Dna, che poi viene integrato nel cromosoma ospite, anche questo, come la Rna polimerasi Rna dipendente, è un enzima piuttosto impreciso: ha lo stesso valore per quanto riguarda gli errori durante il funzionamento, un errore ogni 10^3 - 10⁶ basi aggiunte. Il genoma dell'HIV, che nel corso della propria replicazione utilizza una trascrittasi inversa con questo tasso mutazionale, ha un genoma costituito da 10⁴ ribonucleotidi. Ogni volta che la trascrittasi inversa legge, o meglio copia questo genoma a Rna per formare una molecola di Dna, inserisce per lo meno un errore. Se noi pensiamo che in un individuo infetto da HIV, si può arrivare ad avere la produzione di 10^11 particelle di HIV ci rendiamo conto dei mutanti che insorgono in un solo ciclo di replicazione nelle 24 ore. Questo giustifica la difficoltà ad ottenere un vaccino, lo stesso è vero per un farmaco.
Non è solo importante per l'insorgenza e per i risvolti di una malattia, il funzionamento delle polimerasi ma: il vaccino antipolio, ne abbiamo a disposizione due, il vaccino di Salk con il virus inattivato e il vaccino di Sabin che è invece un virus attenuato ossia un virus vivo che ha perso il proprio potere patogeno. Quando ad un bambino sano viene somministrato questo vaccino di Sabin abbiamo che il virus attenuato replica nell'intestino di questo bambino, suscita quindi una risposta anticorpale efficace nel proteggere quel bambino quando incontrerà il wild type, ossia il ceppo selvaggio della malattia, però il sistema immunitario di questo bambino è valido e quindi nell'arco di un mese, un mese e mezzo, elimina il principio vaccinante, il virus attenuato ha stabilito una bella immunità e tutto è andato liscio.
Cosa succede se questo vaccino viene somministrato ad un bambino molto piccolo che presenta problemi al sistema immunitario con immuno-deficienza?
In questo bambino il sistema immunitario non è capace di debellare il virus, il virus attenuato continua a replicarsi. Questo cosa comporta? Il Poliovirus è un virus a Rna e affida la propria replicazione ad una Rna polimerasi Rna dipendente estremamente inaccurata, si allunga il periodo di sopravivenza del virus nel bambino vaccinato, aumenta la possibilità che a seguito di errori della polimerasi il ceppo che ha perso la propria virulenza, abbia, per mutazione la probabilità di ritornare ad essere nuovamente virulento.
Continua a leggere:
- Successivo: Virus e malattie umane
- Precedente: Che cos’è un virus?
Dettagli appunto:
-
Autore:
Simone Pisu
[Visita la sua tesi: "Caratterizzazione molecolare ed eterogenicità delle varianti emoglobiniche in Sardegna"]
- Università: Università degli Studi di Cagliari
- Facoltà: Biologia
- Corso: Biologia Cellulare e Molecolare
- Esame: Virologia molecolare
- Docente: Prof. Marongiu
Altri appunti correlati:
- Microbiologia clinica
- Biologia applicata
- Microbiologia
- Microbiologia Medica
- Filologia della letteratura italiana
Per approfondire questo argomento, consulta le Tesi:
- Piante medicinali come ausilio per contrastare le patologie virali: tre casi-studio
- Contributo allo studio ed alla terapia dell'epatite C nel paziente HIV-positivo
- Memetica e nuovi media
- Peptidi overlapping derivanti da sequenze gH-omologhe di HSV-1 come inibitori dell'attività virale.
- Selezione di Mutanti HIV-1 Resistenti e Basi Molecolari della Resistenza di Nuovi Inibitori Non-Nucleosidici della Trascrittasi Inversa
Puoi scaricare gratuitamente questo appunto in versione integrale.
Forse potrebbe interessarti:
Filologia della letteratura italiana
PAROLE CHIAVE:
virusgenoma
uncoating
particelle virali
dna genomico
retrovirus
deossiribo virus
ribovirus
 Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Login
Oppure utilizza il tuo account
 o
o
