
Ovaio
È una ghiandola endocrina. Ci sono due ormoni ipofisari che stimolano le gonadi: FSH (ormone follicolostimolante) e LH (ormone luteinizzante), anche chiamati "gonado-tropine ipofisarie". Invece nell'uomo ci sono le cellule di Leydig (producono testosterone). L'ovaio si trova nella piccola pelvi, cioè la parte più bassa (porzione più "declive"), si trova nella parte inferiore della cavità addomino- pelvica (cavità pelvica: grande pelvi e piccola pelvi); nella pelvi trovo: utero, prostata, vescica, tube, ovaie, intestino retto (organi pelvici).
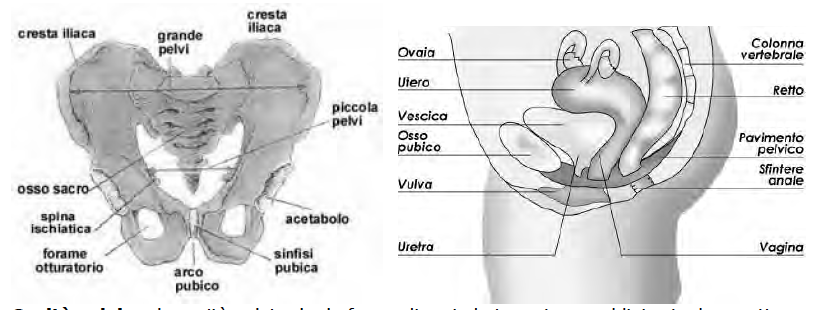
Cavità pelvica: la cavità pelvica ha la forma di un imbuto e viene suddivisa in due parti, una superiore, detta grande pelvi, ed una inferiore, detta piccola pelvi. La grande pelvi fa parte della cavità addominale, la piccola pelvi è la cavità pelvica.
L'ovaio (organo pari) ha la forma di una "mandorla" lunga circa 40 mm. Ha notevole consistenza e la sua superficie è liscia prima della pubertà, mentre durante il periodo fertile diventa irregolare per la presenza di follicoli maturi, di corpi lutei e di cicatrici connettivali.
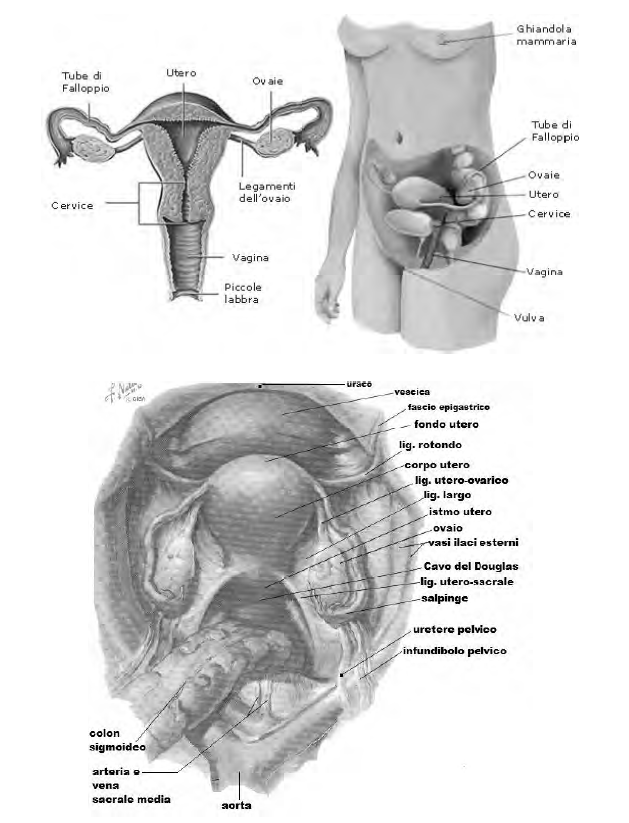
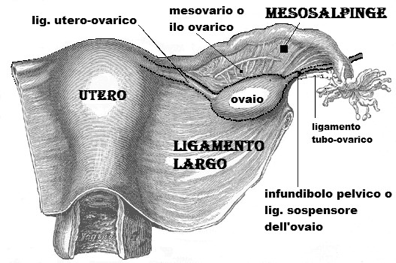
La faccia mediale dell'ovaio volge verso l'utero ed è in parte coperta dalla tuba uterina; la faccia laterale è a ridosso della parete della piccola pelvi e del muscolo otturatorio interno; il margine anteriore è in vicinanza dell'inserzione laterale del legamento largo dell'utero, cui è collegato mediante il mesovario e presenta l'ilo dell'ovaio; il margine posteriore è libero; il polo superiore, detto tubarico, entra in contatto con l'infundibolo della tuba uterina; il polo inferiore, uterino, è collegato all'utero per mezzo del legamento uterovarico.
L'ovaio è posto anteriormente e inferiormente alla biforcazione dei vasi iliaci comuni e occupa la fossetta ovarica. Quest'ultima è una piccola depressione delimitata in avanti dall'inserzione del legamento largo alla parete della piccola pelvi; in alto dai vasi iliaci esterni, posteriormente dell'uretere, inferiormente dall'origine delle arterie uterina e vescicale superiore.
L'ovaio è fissato nella sua posizione da quattro legamenti: il legamento sospensore è organizzato intorno ai vasi ovarici e raggiunge il polo superiore dell'organo; il legamento uterovarico decorre nello spessore del legamento largo e unisce l'angolo supero-laterale dell'utero al polo inferiore dell'ovaio; il mesovario è formato dal peritoneo del legamento largo e unisce quest'ultimo al margine anteriore dell'ovaio; il legamento tubovarico è una piccola piega che unisce il polo superiore dell'ovaio a una fimbria del padiglione della tuba uterina.
La borsa ovarica è una piccola tasca, dipendenza del peritoneo, posta tra la faccia mediale dell'ovaio e la lamina peritoneale che raggiunge la tuba uterina (mesosalpinge), e tra la faccia laterale dell'ovaio e la parete laterale della piccola pelvi; ha il compito di accogliere gli oociti rilasciati dall'ovaio prima che essi vengano aspirati nella tuba.
Struttura dell'ovaio
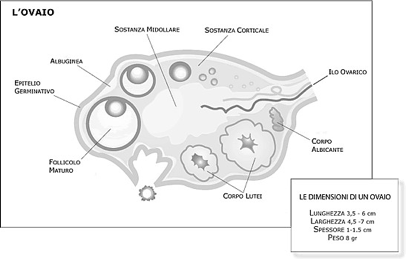
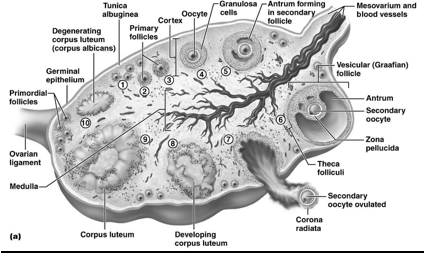
Nel parenchima di un ovaio riconosco una struttura periferica "corticale", che si distingue dalla "midollare".
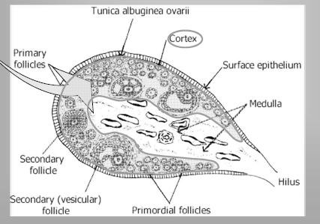
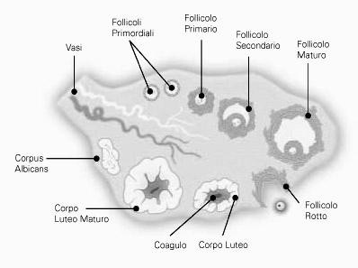
La zona corticale è la parte più periferica dell'organo ed è caratterizzata dalla presenza dei follicoli oofori in vari stadi di maturazione e dalle formazioni che da essi derivano: i corpi lutei, i follicoli atresici e i corpi albicanti. I follicoli oofori sono immersi in uno stroma di tessuto connettivo, ricco di cellule, che partecipa alle modificazioni morfologiche e funzionali dei follicoli stessi durante il ciclo ovarico. I follicoli oofori possono essere distinti in primordiali, primari, secondari, antrali, maturi e atresici. Nello stroma connettivale sono contenuti anche cordoni o gruppi di cellule endocrine (ghiandola interstiziale dell'ovaio), che derivano dai follicoli atresici e producono testosterone e estrogeni.
Nella zona midollare ci sono vasi sanguigni che entrano nell'organo tramite l'ilo ovarico. Le arterie ovariche provengono dall'aorta addominale, tra i rami pari dell'aorta addominale ci sono le arterie gonadiche.
All'interno di un ovaio ci sono numerosi follicoli ovarici; in un ovaio al momento della nascita i follicoli sono già presenti tutti, invece nell'uomo gli spermatozoi si rigenerano continuamente. La mestruazione è l'evento finale di un processo durato un mese che non ha visto la fecondazione della cellula uovo.
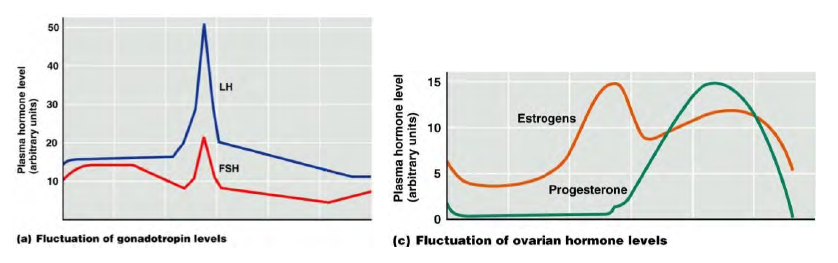
Ciclo ovarico: la prima spinta di maturazione del follicolo è data dall'FSH (fase follicolare), la seconda parte del ciclo è regolata dall'LH.
1. FSH porta alla produzione di estrogeni (E2); via via aumentano gli estrogeni e così essi vanno ad esercitare un feedback negativo sull'ipofisi: l'FSH si riduce.
2. Quando E2 supera un certo livello non fa più da freno, ma da stimolazione dell'ipofisi: aumenta la produzione di LH (picco di LH a metà ciclo) → stimola le cellule ovariche a produrre il progesterone → il progesterone aumenta → giunto a un certo livello, esercita un feedback negativo → se non c'è la fecondazione, niente mantiene la produzione di P e tutto decade → distacco dell'ovulo e dell'endometrio col ciclo mestruale e comincia un ciclo ovarico successivo. Se invece c'è la fecondazione: Beta-Hcg (simile a LH) mantiene P; verificare la presenza di Beta- Hcg è l'esame per vedere a che punto è la gravidanza. Beta-Hcg è prodotta dal trofoblasto (un tessuto che con il procedere della gravidanza si trasformerà successivamente nella placenta), tiene alto il progesterone e dall'analisi del sangue si vede la settimana di gravidanza.
Funzioni del progesterone: mantenere la muscolatura uterina liscia e rilassata in modo da permettere un buon atticchimento; lavorare in sinergia con gli estrogeni per stimolare le pareti dell'utero ad aumentare il loro sviluppo; aiutare la formazione della placenta e a concorrere alla nutrizione del bambino; rallentare le attività dell'intestino in modo da assorbire più sostanze nutritive.
Evoluzione dei follicoli oofori:
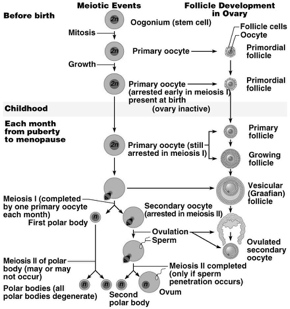
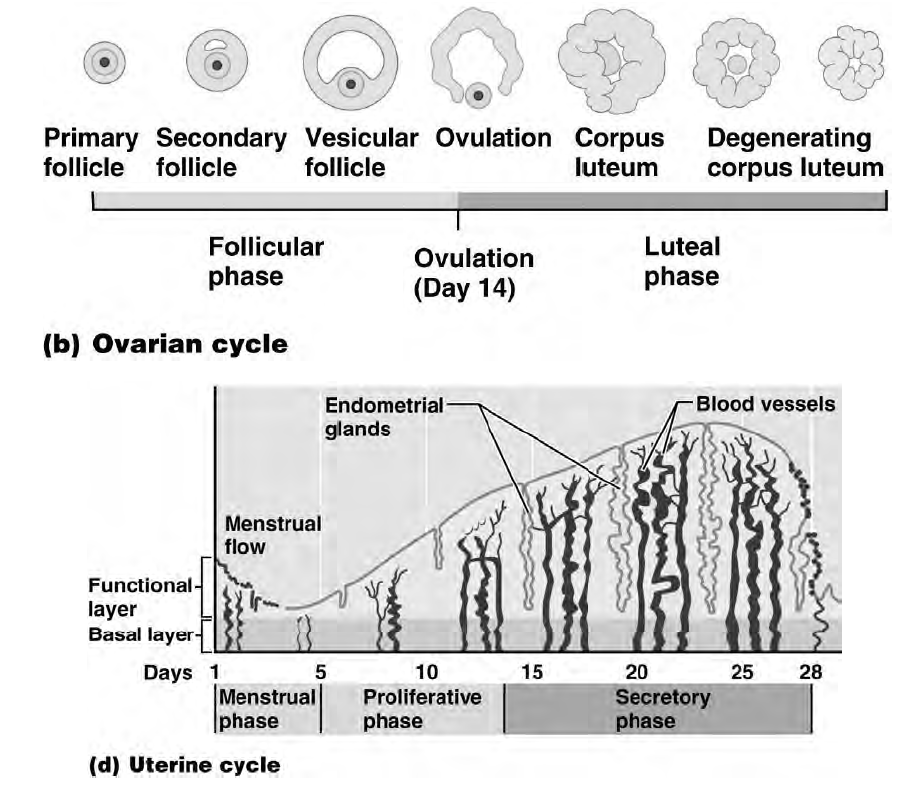
I follicoli oofori maturano per un complesso processo di trasformazione che, partendo dal follicolo primordiale, attraverso le fasi successive di follicolo primario e secondario, porta alla fomazione degli involucri tecali, all'acquisizione della cavità antrale (follicolo antrale o vescicoloso di Graaf), fino alla formazione di un follicolo maturo di grandi dimensioni (19-24 mm di diametro), pronto per il rilascio dell'oocito maturo (ovulazione). Parallelamente all'evoluzione del follicolo ooforo, e sotto il controllo metabolico dello stesso follicolo, l'oocito in esso contenuto segue un peculiare processo maturativo caratterizzato dal progredire della meiosi.
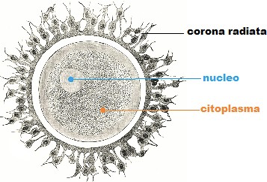
L'ovocito è circondato da epitelio follicolare, che è stimolato dall'FSH, così il follicolo aumenta di dimensioni, all'interno si forma la cavità follicolare in cui è raccolto il liquor follicoli; la cellula uovo assume una posizione periferica. Il follicolo diventa ancora più grosso: follicolo maturo di Graaf, con ovocita prossimo alla superficie, esattamente nel punto in cui avverrà la rottura; infatti poi abbiamo la rottura del follicolo per lisi enzimatica: vengono rilasciati enzimi che permettono alla parete del follicolo, sopra la cellula uovo, di rompersi e espellere l'ovocita. La cellula uovo è espulsa nella tuba uterina. La cellula uovo è circondata dalla corona radiata (strato di cellule follicolari che si porta con sé l'ovocita quando esce dal follicolo).
Le cellule che circondano un follicolo che sta maturando sono le cellule della teca (teca interna e teca esterna). Le cellule della teca interna producono androgeni. La teca esterna sono elementi connettivali che non hanno un ruolo endocrino.
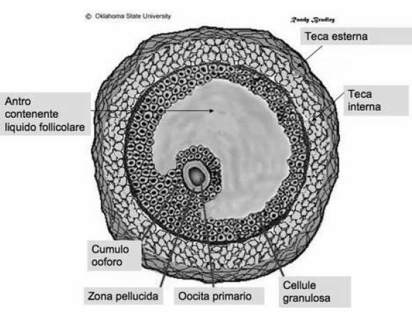
La parete più interna del follicolo è costituita dalle cellule della granulosa, le quali prendono gli androgeni e li convertono in estrogeni e progesterone, tramite un enzima che si chiama "aromatasi" (rompe l'anello aromatico). Gli estrogeni e il progesterone servono a modificare la parete interna dell'utero (il loro bersaglio è la mucosa uterina, l'endometrio, affinché essa subisca modificazione funzionali all'eventuale accoglimento della cellula uovo fecondata).
La zona pellucida è una membrana glicoproteica che avvolge l'uovo nei mammiferi. In particolare si tratta di un involucro primario (prodotto dall'oocita stesso durante l'oogenesi) posto tra la membrana plasmatica della cellula uovo e le cellule follicolari. Contiene i recettori per gli spermatozoi e contribuisce al blocco della polispermia. Inoltre serve a proteggere l’embrione durante la segmentazione, induce la liberazione degli enzimi litici presenti nell'acrosoma e garantisce la specie-specificità della fecondazione.
Gli spermatozoi sono prodotti nel didimo e poi sono soggetti al processo di capacitazione nel epididimio; quando la cellula uovo è stata fecondata cambia i profili recettoriali: nessuno spermatozoo può più entrare.
Fecondazione non è sinonimo di gravidanza, perché magari una cellula uovo è fecondata ma poi quando arriva nell'utero non trova una zona adatta per annidarsi.
Il progesterone è importante per la seconda metà del ciclo (importante per una eventuale gestazione).
Ciò che rimane del follicolo dopo che è stata espulsa la cellula uovo (quindi dopo l'ovulazione) si chiama corpo luteo. Esso ha il ruolo di produrre progesterone. Il destino del corpo luteo dipende dal fatto se c'è stato o meno l'annidamento:
1. Nella maggior parte dei casi diventa corpo luteo mestruale: se non c'è stato l'annidamento i livelli di progesterone aumentano, sostenuti dall'LH (è il picco di LH che induce la rottura del follicolo), fino a che l'LH non arriva a un certo livello per cui l'adenoipofisi smette di produrlo, quindi c'è la caduta del progesterone e avviene la mestruazione, non è solo sanguinamento ma anche desquamazione (tutto ciò che avevamo costruito non serve più, va spazzato via). Quindi, cade il progesterone, le arterie dell'utero si contraggono sempre di più fino a che non avviene una necrosi dell'endometrio (perché il periodo di contrazione è sempre più lungo, quindi arriva sempre meno sangue al tessuto e esso muore).
2. Può diventare un corpo luteo gravideo, cioè deve sostenere la gravidanza; quando l'LH arriva a un certo livello non viene più prodotto, quindi serve qualcosa per far continuare il corpo luteo a produrre progesterone: esistono le gonadotropine corioniche (β-HCG) che stimolano il corpo luteo a produrre progesterone (il corpo luteo le riconosce come LH anche se non lo sono); il corion è la struttura che si è venuta a creare intorno alla cellula uovo fecondata e annidata. Quando si forma la placenta le beta HCG spariscono; esse sono dosate nei test di gravidanza.
Continua a leggere:
- Successivo: Vie genitali
- Precedente: Apparato genitale femminile
Dettagli appunto:
- Autore: Andrea Panepinto
- Università: Università degli Studi di Pisa
- Facoltà: Medicina e Chirurgia
- Corso: Logopedia
- Esame: Anatomia
- Docente: Ruffoli
Altri appunti correlati:
- Anatomia umana
- Fondamenti Anatomofisiologici dell'Attività Psichica
- Lo sviluppo dell'apparato digerente nel feto
- Apparato respiratorio
- L'apparato digerente
Per approfondire questo argomento, consulta le Tesi:
- Lo squilibrio idroelettrolitico e l'importanza dell'alimentazione nel paziente ileostomizzato
- Alla scoperta del sistema nervoso autonomo: La Teoria Polivagale delle emozioni
- L'analisi posturale e la pedana stabilometrica-propriocettiva
- Il Biomeccanico: una nuova figura professionale al servizio del calcio
- Il ruolo del sistema nervoso nell'ipertrofia muscolare
Puoi scaricare gratuitamente questo appunto in versione integrale.
 Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.